Biotische Faktoren
Damit sind alle Wechselbeziehungen einer Art mit dem belebten Teil ihrer Umwelt gemeint, also mit Individuen der eigenen oder einer anderen Art. Das Verständnis dieser Beziehungen gibt direkte Aufschlüsse über die Populationsentwicklung, und - bezogen auf die interspezifischen Beziehungen - auch darüber, ob sich eine Art überhaupt langfristig in einem Ökosystem etablieren kann, etwa im Lichte des Konkurrenzausschluss-Prinzipes betrachtet.Interspezifische Beziehungen (Symbiosen)
Die Wechselbeziehungen zu Individuen anderer Arten. Nach dem Konzept von Warder Clyde Allee (1885 ‐ 1955, US-amerikanischer Zoologe und Ökologe) lassen sich interspezifische Wechselwirkungen zweier Arten nach einem Schaden-Nutzen-Schema klassifizieren, wobei in der nun folgenden Darstellung aber nur direkte Interaktionen beschrieben werden, ohne dabei langfristige Konsequenzen in Betracht zu ziehen. Was damit gemeint ist, wird weiter unten erklärt.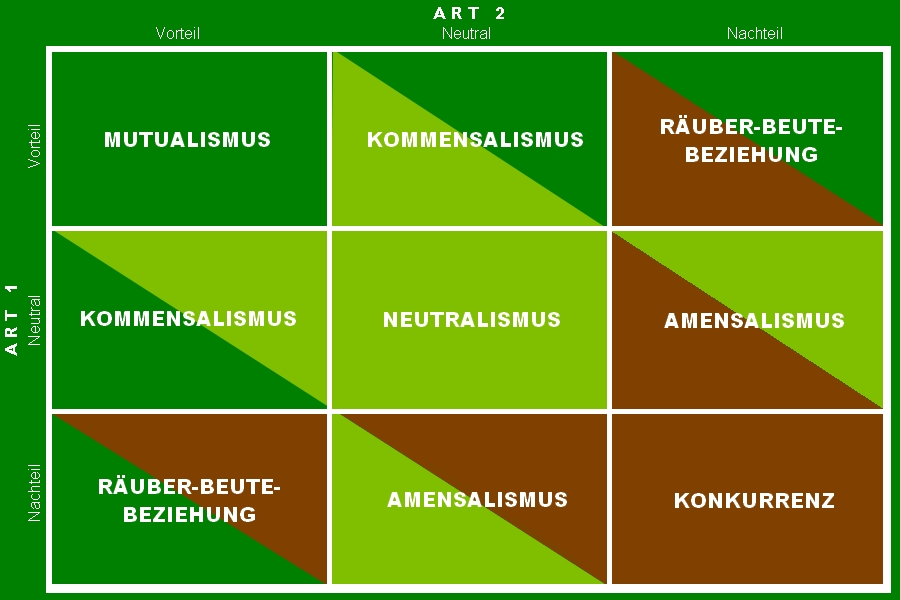
Abb. 1: Interspezifische Beziehungen zwischen zwei Arten.
Mutalismus (+/+)
M. liegt immer dann vor, wenn beide Arten durch ihre Wechselbeziehung einen klaren Vorteil haben. Die Populationsdichte einer Arten wird daher durch das Auftreten der anderen Art günstig beeinflusst. Es kann weiter danach differenziert werden, inwieweit sich eine Abhängigkeit entwickelt hat, also ob der Mutualismus fakultativ oder obligatorisch ist. Außerdem kann auch danach differenziert werden, ob eine oder beide Arten durch andere ersetzbar sind, oder nicht. Daraus ergeben sich drei Möglichkeiten:Tabelle 1: Formen von Mutualismus.
Allianz |
Kooperation |
Eusymbiose |
| Mutualismus nicht in beiden Spezies zwingend artspezifisch, Interaktion ist substituierbar. | Mutualismus ist zwar artspezifisch, jedoch nicht beiderseitig essentiell. | Mutualismus ist streng artspezifisch und für beide Partner essentiell - es hat sich eine wechselseitige Abhängigkeit voneinander ergeben. |
| Beispiele: | ||
| Die meisten Blütenpflanzen werden von Insekten bestäubt, wobei weder die Pflanze auf ein bestimmtes Insekt, und umgekehrt das Insekt nicht auf eine bestimmte Pflanze spezialisiert ist. Es handelt sich also um polyphile Pflanzen. | Fabaceen leben mit Rhizobien in einer Symbiose. Es ist aber möglich, dass einerseits die Rhizobien auch außerhalb einer Pflanze lebensfähig sind, andererseits können auch Fabaceen ohne Rhizobien gedeihen - sind dann aber auf nährstoffreichere Standorte angewiesen. | Mykorrhiza der Orchideen: Hier symbiontieren eine Pilzart mit einer jeweils einer Orchideenart. Weder Pilz noch Orchidee sind ohne den Partner lebensfähig. Solche Pflanzen sind monophil. |
Ausprägung und Lokalisation: Dringt der Symbiont in seinen Wirt ein, so wird von Endosymbiose gesprochen. Hierfür ist die Mykorrhiza der Landpflanzen (Embryophyta) mit verschiedenen Pilzen das bekannteste Beispiel. Agieren Symbiont und Wirt aber räumlich eigenständig, so wird von einer Exosymbiose gesprochen.
Kommensalismus oder Probiose (+/0)
Kommensalismus (lat. con = mit, mensa = Tisch) bezeichnete ursprünglich ein Verhältnis zweier Individuen zueinander, bei der eines sich vom anderen ernährt, ihm dabei aber im Gegensatz zum Parasitismus keinen Nachteil brachte. Allen hat den Begriff dann verallgemeinert und erweitert, und beschrieb damit alle Wechselbeziehungen zwischen zwei verschiedenartlichen Individuen, die nur einem der beiden vorteilhaft erscheinen, dem anderen jedoch auch nicht schaden. Evolutionär kann Kommensalismus aus Mutualismus entstehen, und umgekehrt kann Kommensalismus zu Mutualismus oder zu einer Räuber-Beute-Beziehung (Dualismus) werden.Tabelle 2: Formen von Kommensalismus.
PhoresieLogistische Interaktion |
SaprophagieTrophische Interaktion |
SymphorismusRäumliche Interaktion |
| Phoret nutzt einen Phorenten zur Ortsveränderung. | Kommunsale nutzt Nahrungsrückstand. | Besiedlungs-Interaktion einer Art 1 in Abhängigkeit von Art 2. |
|
1. Epiphilie und Epichorie:
Bestäubung bzw. Samenverbreitung durch Anheften an Tiere, wobei in erstem Falle kein Vorteil (etwa durch Bereitstellen von Nektar) gewährt wird. 2. Eigentliche Phoresie: Ein Tier lässt sich von einem anderen transportieren. 3. Hyperphoresie: Ein Tier, das seinerseits zum eigenen Transport an ein anderes Tier angeheftet ist, ist selber Träger dritter Tiere, die sich ihrerseits von ihm transportieren lassen. |
1. Nekrophagie / Zoosaprophagie: Ernährung von totem tierischen Material, Aasfresser. 2. Koprophagie: Ernährung von tierischen Exkrementen. 3. Phytosaprophagie: Ernährung von totem pflanzlichen Material. 4. Nekromenie: Ernährung von Überresten eines Organismusses nach dessen Tod, wobei dieser bereits zu Lebzeiten besiedelt wird. Der sich ernährende Organismus trägt nicht zu dessen Tod bei (Ggs. Parasitoismus). |
1. Eigentl. Symphorismus oder Epökie: Aufsiedelung eines Tieres (Epizoe) oder einer Pflanze (Epiphyt) auf einer anderen, ohne dieser zu schaden. 2. Synökie / "Wohngemeinschaft": Eine Art nutzt zusammen mit einer anderen Art gemeinsamen Wohnraum. In der Regel ist der Mitbewohner dabei wesentlich kleiner als der Hauptbewohner, und beeinträchtigt nicht dessen Futterquelle. 3. Parökie / "Beisiedelung": Nachbarschaftliches Besiedeln einer Art neben einer anderen, die dieser entweder indirekt Futter verschafft oder Schutz bietet. 4. Entökie / "Einnistung": Einnistung eines kleinen Organismus in einen größeren, ohne ihn zu parasitieren. 5. Metabiose: Eine Art 1 schafft durch ihre Tätigkeiten oder Lebensstil die Bedingungen dafür, dass eine Art 2 leben kann. |
Neutralismus oder Gleichgültigkeit (0/0)
Bringt eine Wechselwirkung zwischen Art 1 und Art 2 keiner der beiden einen Vorteil oder Nachteil, so wird von Neutralismus gesprochen. Der Charakter einer neutralen Interaktion ist oftmals nur schwer nachweisbar, da Vor- oder Nachteile nicht immer direkt ersichtlich sind.Beispiel: Die Besiedlung einer Miesmuschel mit Seepocken ist für beide Tiere weder schädlich noch nützlich.
Amensalismus (−/0)
Als Amensalismus wird eine Interaktion zwischen zwei Arten bezeichnet, die einer einen Nachteil bringt, der anderen aber weder nützt noch schadet. Im Gegensatz zu anderen Interaktionen ist Amensalismus oft nur ein Produkt des Zufalls, ohne dass die agierende Spezies eine Absicht damit verfolgt.Tabelle 3: Formen von Amensalismus.
Faktorischer AmensalismusSchädigung durch Verdrängen |
Physischer AmensalismusDirekte physische Schädigung |
|
| Art 1 verdrängt Art 2 von einer Ressource ohne mit ihr um diese zu konkurrieren, oder ohne sie selber zu benötigen. | 1. Exogene Schädigung: Art 1 beeinträchtigt Art 2 durch direkte physische Gewalteinwirkung, ohne dabei eine Verdrängung oder Nahrungserwerb zu beabsichtigen. |
2. Endogene Schädigung: Art 1 beeinträchtigt Art 2 durch ein Toxin, ohne dabei Verdrängung oder Nahrungserwerb zu beabsichtigen. |
| Beispiele: | ||
| Ein bereits großer Baum überwächst eine sehr kleine Pflanze und entzieht ihr dadurch das Licht. (Hier besteht ein Übergang zu Konkurrenz) | Eine Herde von Rindern zertrampelt Pflanzen, ohne diese abweiden zu wollen. | Ein Pilz produziert durch Stoffwechsel einen Giftstoff der andere Organismen zu schädigen vermag, ohne dass dies dem Pilz einen Vorteil verschafft. |
Konkurrenz (−/−)
Als Konkurrenz wird eine biotische Interaktion um Ressourcen bezeichnet, die für beide Beteiligten nachteilig ist. Konkurrenz ist ein evolutiver Selektionsfaktor!
Abb. 2: Formen der Konkurrenz
Tabelle 4: Formen der Konkurrenz
| Exploitative Konkurrenz Beeinträchtigungskonkurrenz oder auch indirekte/unvollständige Konkurrenz: Ressourcen werden verbraucht, und stehen daher dem Kontrahenten nicht mehr zur Verfügung. |
Interferenz-Konkurrenz Verdrängungskonkurrenz oder auch direkte Konkurrenz: Ein Kontrahent verdrängt den anderen von einer Ressource durch direkte Interaktion (z. B. Kämpfe). |
||
| inter- | spezi-fisch |
Beispiel: Wölfe und Luchse konkurrieren in einem Habitat um den Reh-Bestand miteinander. | Beispiel: Nahrungskonkurrenz an einem Vogelhaus, wobei beispielsweise Rotkehlchen Blaumeisen vertreiben können. |
| intra- | Beispiel: Nahrungskonkurrenz juveniler Säugetiere oder Vögel im Nest/Bau. Diese kann so weit gehen, dass die schwächeren Jungen verhungern. | Beispiel: Intrasexuelle Selektion um Paarungspartner stellt eine Verdrängung unterlegener Artgenossen durch fittere dar, und kann daher auch als intraspezifische Konkurrenz gesehen werden (Ressource: Paarungspartner). |
Einige Hypothesen und Theorien der Ökologie, die mit Konkurrenz zusammenhängen:
Konkurrenz-Ausschluss-Prinzip
Dieses besagt, dass zwei Arten nicht die exakt gleiche ökologische Nische besetzen können, ohne dass eine der beiden Arten verdrängt werden würde. Dieses Prinzip widerlegt auch die laienhafte Ansicht, wonach "in der Natur keine Spezies eine andere ausrotten könne."Die Hypothese berücksichtigt allerdings keine Störungen oder umweltbedingte Stressfaktoren: Durch diese - in der Realität permanent oder temporär auftretenden Einflüsse - kann Verdrängung verhindert oder zeitlich weit hinausgezögert werden. Verdrängung findet nur dann statt, wenn die Zeit zwischen Störungsereignissen länger ist, als dass die konkurrenzstärkere Art bräuchte, um die konkurrenzschwächere vollständig zu verdrängen.
Hypothese der stabilen Koexistenz (Lotka-Volterra-Modell)
Stabile Koexistenz zweier Arten sei nur dann möglich, wenn deren intraspezifische Konkurrenz jeweils größer ist, als die interspezifische.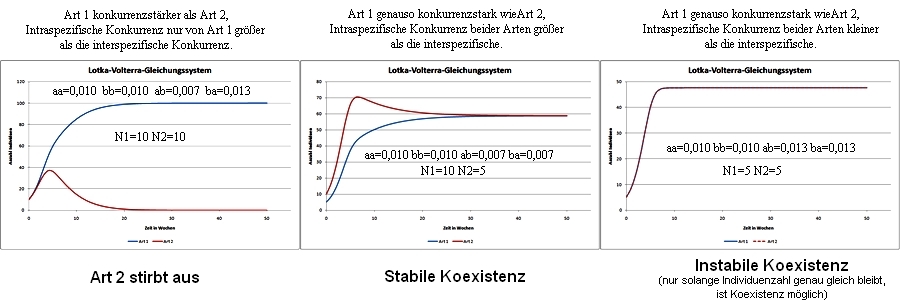
Abb. 3: Populationsentwicklung für verschiedene Szenarien. aa, bb, ab, ba = Konkurrenzvariablen (aa,bb = intraspezifisch); N = Individuenzahl.
Diese Hypothese ist sehr theoretisch, da sie einige Aspekte der realen Umwelt außer Acht lässt: Es wird davon ausgegangen, dass die Wechselwirkung absolut sei, sich also die Konkurrenten nicht aus dem Weg gehen könnten, so wie es in der Natur oft beobachtet werden kann. Störungen und Stress können die Populationsentwicklung ebenfalls beeinflussen, sodass trotz ungünstiger Konkurrenzbedingungen dennoch eine Koexistenz möglich ist. Daraus lässt sich die nächste Theorie herleiten:
Theorie der mittleren Störungshäufigkeit
|
Aus diesen beiden Hypothesen kann gefolgert werden, dass die Artenzahl in einem Biotop dann am höchsten ist, wenn es einer mittleren Störungsfrequenz unterworfen ist:
Sind die Störungen zu selten, können konkurrenzschwächere Arten vollständig von den konkurrenzstärkeren verdrängt werden (nur einige wenige K-Strategen koexistieren). Sind die Störungsereignisse dagegen zu häufig, so werden nur einige wenige stressresistente r-Strategen koexistieren können, wodurch die Artenzahl ebenfalls niedrig sein wird. Beispiele für beide Extreme: Umwelten mit seltenen Störungen lassen in Mitteleuropa Rotbuchenwälder entstehen, in denen einige K-Strategen unter hohem Konkurrenzdruck koexistieren. Ein Beispiel sei der Waldmeister-Rotbuchenwald, der überaus artenarm ist. Das Gegenbeispiel kann an Ufern naturbelassener Flussläufe vorgefunden werden, wo durch Überschwemmungen und Eisgang jährlich Störungen stattfinden, und sich daher fast nur einige wenige r-Strategen bzw. Pionierpflanzen ansiedeln können (z. B. Bruchweide, Salix fragilis). |
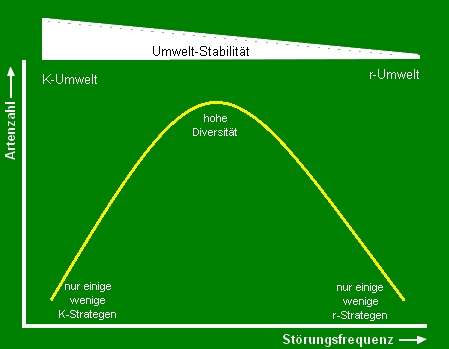 Abb. 4: Auftragung Artenzahl gegen Störungshäufigkeit. |
Trophische Beziehungen = Räuber-Beute-Beziehungen (−/+)
Diese Art der biotischen Interaktion bringt für einen Beteiligten einen Vorteil, den anderen einen Nachteil. Es gibt verschiedene Modelle, wie trophische Beziehungen charakterisiert werden können. Hier wird das Modell nach Allen beschrieben, der trophische Beziehungen nach Intimität (wie viele Organismen werden vom Prädatoren befallen) und Mortalität (ist die Prädation tödlich) unterscheidet:Tabelle 5: Trophische Beziehungen
| Mortalität ist niedrig | Mortalität ist hoch | |
| Intimität ist niedrig | WEIDEGÄNGER | ECHTER RÄUBER |
| Intimität ist hoch | PARASIT | PARASITOID |
Weidegänger
Zu den Weidegängern werden alle Organismen gezählt, die sich zwar von anderen ernähren, sie dabei aber in der Regel nicht töten. Als weiteres Kriterium wird gesehen, dass der Weidegänger im Laufe seines eigenen Lebens eine Vielzahl von Organismen zwecks Ernährung befällt. Beispiele für Weidegänger: Kühe, die Gräser abweiden. Die einzelnen Pflanzen werden in der Regel nicht getötet, sondern sind in der Lage, nochmals auszutreiben (bodennahe Interkalarmeristeme als Anpassung an Weidegänger). Ein weiteres Beispiel sind Zecken, die landläufig unrichtig als "Parasiten" bezeichnet werden: Auch sie befallen ihre Wirtsorganismen dergestalt, dass es im Laufe ihres Lebens (Zecken werden mehrere Jahre alt) viele sind, und diese nicht töten.Echte Räuber
Als Echte Räuber werden Organismen bezeichnet, die im Laufe ihres eigenen Lebens viele Beutetiere töten und verspeisen. Definitionsgemäß ist der Löwe genauso ein Echter Räuber wie der Sonnentau oder die Venus-Fliegenfalle (Familie Droseraceae).Parasiten
Parasiten ernähren sich im Laufe ihres eigenen Lebens nur von einem einzigen Wirt, oder wenigstens nicht vielen, ohne sie dabei jedoch zu töten. Zu ihnen gehören die Kopfläuse, oder beispielsweise Kohlweißlingsraupen, sofern sie ihre Wirtspflanzen nicht töten, sondern nur nicht mortal schädigen. Unter den Pflanzen kann man Vertreter der Orobanchaceae als Parasiten bezeichnen: Sie befallen die Wurzeln von Poaceaeen, und saugen Xylemsaft. Die befallenen Gräser werden dabei nicht getötet.Parasitoide
Parasitoide befallen einen einzigen Wirt und töten ihn nach einer gewissen Zeit ab. Das klassische Beispiel aus der Zoologie sind manche Wespenarten, die Schmetterlingsraupen parasitoitieren: Die Wespe legt dabei ein Ei einer Schmetterlingsraupe ab, die daraufin ganz normal weiter frisst, sich häutet und irgendwann auch verpuppt. Beim Eintritt in das Puppenstadium wird dann die Wespenlarve aktiv und beginnt, die Raupe von innen heraus aufzufressen. Die Wespenlarve verpuppt sich in der toten Raupe und schlüpft als fertig entwickelter Imago. Im Pflanzenreich kann man die Würgefeige (Ficus spec.) als Beispiel für Parasitoismus sehen: Sie befällt Bäume, um sich an ihnen hoch zu ranken, und eigene Biomasse aufzubauen. Schlussendlich tötet sie den Wirtsorganismus, und nimmt gänzlich seinen Platz ein, und ernährt sich von den Überresten desselben.Der Prädationszyklus
| Dieses Modell entstammt der Tier-Ökologie, und beschreibt die verschiedenen Schritte im Beute-Erwerb eines Prädators, und davon abgeleitet auch Strategien der Beute, sich dem Zugriff zu entziehen. Es wird von fünf Schritten ausgegangen, die jeder Räuber durchläuft, wenn er
Beute zum Nahrungserwerb macht: Ist die Verdauung abgeschlossen, so stellt sich ein Hungergefühl ein, das den Räuber dazu bewegt, nach Beute zu suchen. Begegnet er einem Organismus, so muss dieser auch als Beute erkannt werden. Ist dies der Fall, so geht der Räuber in den Angriff über, welcher
bei Erfolg dazu führt, dass die Beute erlegt und gefressen wird. Zwischen jedem der beschriebenen fünf Schritte können potenzielle Beutetiere Verteidigungsmaßnahmen entwickeln, um dadurch dem Zugriff zu entgehen.
Step 1-Verteidigung schaltet sich zwischen Fressen und Suchen, Step 3-Verteidigung zwischen Begegnen und Erkennen usw.: Tabelle 6: Verteidigungs-Strategien im Prädationszyklus beschrieben.
|
 Abb. 5: Prädationszyklus und Verteidigungsmaßnahmen (schematisch). |