AnatomieDie Gametophyten der Moose erscheinen entweder lappig (thallos) oder blättrig (folios), haben aber weder echte Blätter noch echte Wurzeln. Der Sporophyt der Tracheophyten (Gefäßpflanzen) wird als Kormus bezeichnet. Ein Kormus lässt sich in die drei Gewebe Wurzel, Spross und Blatt unterteilen, wobei es im Laufe der Evolution zu zahlreichen Spezialisierungen dieser drei Grundgewebe gekommen ist.So sind beispielsweise Blüten, genau wie die Lagen einer Zwiebel oder die Schuppen eines Tannenzapfens, physiologisch betrachtet Blätter! |
 |
Die Wurzel (Radix)
Der untere Teil des Kormus einer Pflanze wird als Wurzel bezeichnet. Eine Wurzel ist, genau wie Spross und Blatt, immer Teil eines Sporophyten, nie der eines Gametophyten. Dies bedeutet also, dass Moose keine Wurzeln haben! Bei Samenpflanzen treibt beim Keimen die Keimwurzel aus, und bohrt sich in den Boden ein (positiver Gravitropismus). Erst dann entfalten sich die Keimblätter (bzw. bei Monokotyledonen: das Keimblatt). Durch die Wurzeln sind Pflanzen an den Ort gebunden, an dem sie aufkeimten; und dieser Umstand bestimmt ihr ganzes Leben sowie das Konkurrenzverhalten bzgl. Nachbarpflanzen (Kampf ums Licht). Im Gegensatz zum Spross können aus Wurzeln keine Blätter austreiben; Seitenwurzeln entstehen - im Gegensatz zu Seitensprossen - endogen, d. h. sie werden durch Zellteilung aus dem Perizykel (Zentralzylinder), von innen heraus gebildet.Aufgaben der Wurzel
- Verankerung der Pflanze, zumeist im Boden. Daneben können Wurzeln aber auch verschiedene Metamorphosen infolge Spezialisierung erfahren. Dazu weiter unten mehr.
- Transport von Wasser und Mineralstoffen aus dem Boden in die Pflanze. Dies geschieht über die Leitbündel im Zentralzylinder. Dabei wird Wasser vom Xylem, Nährstoffe und Mineralien über das Phloem transportiert.
- Bildung von Phytohormonen, die von dort aus ebenfalls in den Rest der Pflanze transportiert werden (wichtigste Gruppen: Auxine, Cytokinine). Auch sekundäre Pflanzenstoffe werden zumeist in den Wurzeln synthetisiert.
- Ort der Symbiose mit Pilzen oder Bakterien: Die Wurzelhaare können mit Pilzhyphen Zell-Zell-Kontakte eingehen, und dadurch Stoffe miteinander austauschen (Mykorrhiza). Dies ist bei allen Landpflanzen der Fall. Einige Pflanzen symbiotieren mit stickstofffixierenden Bakterien, so z. B. die Erle (Alnus) oder die Hülsenfrüchtler (Fabaceae, bei ihnen: Knöllchenbakterien = Rhizobien). Diese sind dann in Wucherungen des Rindengewebes in vergrößerten Parenchymzellen lokalisiert.

Aufbau einer WurzelDie äußerste Schicht einer Wurzel stellt das Abschlussgewebe dar. Im Bereich zwischen Meristemzone und Wurzelhaarzone wird dieses Abschlussgewebe von der Rhizodermis gebildet. Die Rhizodermis besteht aus einer Reihe nichtcutinisierter Zellen. Oberhalb der Wurzelhaarzone stirbt die Rhizodermis ab, aus der darunterliegenden Hypodermis (teilweise verkorkte Zellen) geht die Exodermis als neues Abschlussgewebe hervor.Unterhalb des Abschlussgewebes befindet sich die Wurzelrinde, ein parenchymales Gewebe (daher auch Rindenparenchym genannt). Durch die Wurzelrinde findet der Stoffaustausch zwischen den Abschlussgeweben und den Leitgeweben statt. Hier ist auch der Ort, an welchem die Pilzhyphen bei Mykorrhiza mit den Pflanzenzellen in Kontakt treten. Als innerste Schicht der Wurzelrinde schließt sich die Endodermis an. Sie ist eine Zelllage dick, und die Zellwände dieser Zellen sind stark ausgeprägt, um dadurch den unkontrollierten Übertritt von Wasser bzw. Nährstoffen aus den Leitgeweben zu verhindern. Der Stoffaustausch zu der Wurzelrinde ist über Durchlasszellen möglich. Noch weiter innen befindet sich der Zentralzylinder, der vom Perikambium zur Endodermis abgegrenz ist. Der Zentralzylinder enthält die Leitgewebe (Xylem und Phloem), die je nach Pflanzenart verschieden aufgebaut sind (Stelen, unter Spross genauer beschrieben). |
Abb 1.: Aufbau der Wurzel |
Primäres Wurzelwachstum
Das Wachstum einer Wurzel erfolgt, wie auch bei Blättern und Sprossen, aus Apikalmeristemen heraus. Dieses Meristem wird an der Spitze durch die Calyptra oder Wurzelhaube geschützt. Sie besteht aus einigen Lagen kurzlebiger parenchymatischer Zellen, die zudem schleimbildend sind, und dadurch das Durchdringen des Erdreiches begünstigen.Oberhalb der Calyptra befindet sich die Meristemzone, also der Ort, wo die teilungsfähigen und nicht ausdifferenzierten Meristemzellen (das sind die Stammzellen der Pflanzen) liegen. Diese scheiden nach unten hin ständig Wurzelhaubenzellen ab.
Nach oben hin - in die Streckungs- und Determinierungszone werden sich zu den verschiedenen Gewebetypen ausdifferenzierende Zellen aus dem Meristem heraus gebildet.
Engütige Ausdifferenzierung erlangen die Zellen dann in der Wurzelhaarzone, und nur hier können aus einer Wurzel heraus Wurzelhaare gebildet werden: Bis zur Wurzelhaarzone bildet die Rhizodermis das Abschlussgewebe der Wurzel, und aus ihr heraus können Wurzelhaare gebildet werden. Diese vergrößern die resorptionsfähige Oberfläche der Wurzel enorm, und erhöhen dadurch die Effizienz der Wasser- und Mineralstoffaufnahme. Wurzelhaare haben eine Lebensdauer von nur wenigen Tagen und werden infolge des weiteren Wurzelwachstums daher ständig neu gebildet.
Der oberste Wurzelabschnitt ist die Differenzierungzone. Hier sind alle Gewebe vollständig ausdifferenziert und die Zellen der Rhizodermis abgestorben. Das Abschlussgewebe wird ab hier von der Exodermis gebildet, die aus der ursprünglichen Hypodermis hervorgegangen ist.
Sekundäres Dickenwachstum
(Abschnitt wird fortgesetzt)Der Spross
(Abschnitt muss noch erstellt werden)Das Blatt
(Abschnitt muss noch erstellt werden)Die Blüte
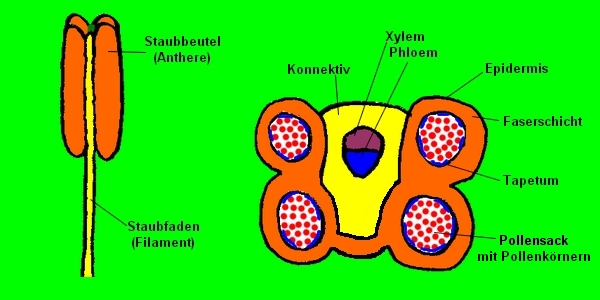
Bei der Blüte handelt es sich physiologisch um differenzierte Blätter, und nicht um einen etwaigen vierten Gewebetyp!